
INFLeXions No. 6 – Arakawa + Gins special issue of Inflexions
Bioscleave: Shaping our Biological Niches by Stanley Shostak, University of Pittsburgh, USA
view PDF version
Go to INFLeXions No. _
6 |5 | 4 | 3 | 2 | 1
focused on the body imbued with the mechanics of life and serving as the basis of the biological processes: propagation, births and mortality, the level of health, life expectancy and longevity, with all the conditions that can cause these to vary (Foucault, 1980: 139).
Fortunately, an artist and a poet have created a shelter from biopolitics in Bioscleave House and provided an inspiration to live in real time. Of course, biopoliticians grumble that claims made for Bioscleave House are unscientific, anecdotal, and lack controls. But scientific studies in public health and disease management are frequently heuristic, beginning with anecdotal evidencewith exploratory studiesand 150,000 years of human evolution have provided all the controls one needs!
In any event, if Bioscleave House were a drug assessed under the protocols of regulatory agencies (e.g., the FDA in the US or EMEA in the EU), it would already have passed Phase 0that human beings process the drug and the drug works in the human being as expected. It would be time to move on to Phase I in earnest! In Phase I, Bioscleave House would be tested on a small number of healthy volunteers to see if objectives are validated by results. Phase II would test Bioscleave House's impact on wellbeing and longevity in a larger number of volunteers drawn from an enlarged pool of possible subjects. Finally, having demonstrated that Bioscleave House works as intended, it would be ready for Phase III, multicenter trials on large groups for long durations aimed at the definitive assessment of effectiveness in comparison with the current "gold standard," namely life as we know itshaped by biopolitics. At this point, Arakawa and Gins would submit applications to the regulatory agencies that would permit volunteers to obtain Bioscleave Houses independently. Finally, during Post Marketing Surveillance Trials (i.e., Phase IV), the label would be expanded to incorporate additional evidence for the Bioscleave House efficacy in individuals not included in the population for which Bioscleave House was originally approved for marketing.
Of course, this scenario would raise hackles among those living by the dictates and standards of biopolitics. Biopolitics supports "anti-aging" medicine whereas Bioscleave House is "pro-aging" without medicine. Aging is a problem for biopolitics but not for Arakawa and Gins. Rather, living fully at every age is the problem they confront. Biopolitics would have increasing numbers of human beings living fragile and vulnerable lives as nonagenarians, centenarians, and supercentenarians. Bioscleave House employs biotopology to extend vigorous life throughout prolonged adulthood. Biopoliticians make metaphysical claims for imminent and permanent cures of disease associated with aging while Bioscleave House espouses human enhancement and the evolution of vigorous life, promoting healthy living now and in generations to come.
The difference between biopolitics and biotopology is easily illustrated.
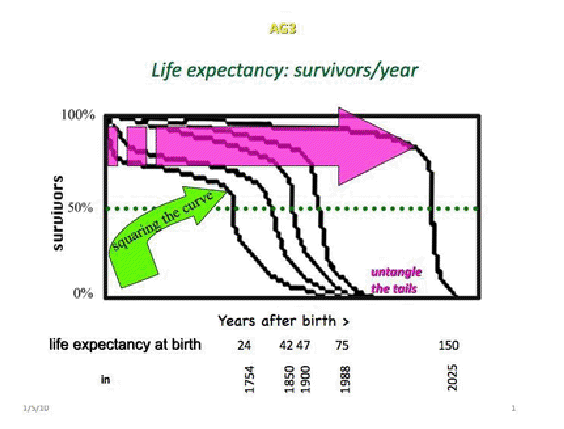
The above image shows five survivorship curves, also known as human life expectancy curves, tracing the percentage of individuals ('survivors') alive in a cohort as they age ('years after birth '). The four curves toward the left are based on data for people in the United States and Europe, actuarial extrapolations, and smoothing algorithms. The one curve at the right is based entirely on projections. The four data-based curves represent cohorts of individuals born respectively in 1754, 1850, 1900, and 1988; the fifth curve is for an entirely hypothetical cohort of individuals to be born in 2025.
The curves all begin at 100%, when all members of the cohort are alive, and end at 0%, when all members of the cohort are dead. A plateau is reached in each curve during adult life followed by a period of rapid decline when survivorship drops off precipitously until moderating and approaching zero asymptotically in old age.
Several important points emerge from seeing the four data-based curves together: The first point is that the four curves follow a similar pattern in which a more or less horizontal arm meets a more or less vertical arm. The second point is that the more or less horizontal arms move upward and lengthen while the more or less vertical arms become increasingly upright. The third point is that the "tails" of the four curves overlap (i.e., are entangled) as they approach 0, at the bottom of the graph. As a result, the shape of the curves changes from somewhat rounder on the left to somewhat squarer on the right. Called "squaring the curve," biopoliticians attribute the effect to improvements in health care management.
Thus, the more horizontal portions of the curves have risen and flattened due to improvements in pre- and post-natal care of women, neonatal care, vaccination, and treatment of infectious diseases among the young. Consequently more babies have survived to become juveniles and more preadolescents have advanced into adulthood.[1] Simultaneously, the more vertical portion of the curve is pushed to the right by the increased numbers of young people surviving into adulthood and by middle-aged people surviving longer. These changes are generally attributed to reductions in exposure to hazards such as those in polluted air, water, and cigarette smoke, and to increased time available to individuals for rest, allowing their bodies to recuperate from the daily assaults of normal life, especially those suffered at work. In addition, survival is promoted by improved treatment of chronic diseasealthough the rampant epidemics of obesity and type II diabetes suggest we are not doing everything we should be doing to combat chronic disease.
How many years have been added to human life as a result of squaring the curve? With a little coaxing this question is answered with numbers generated from these curves. The dotted horizontal line bisecting each curve at 50% (i.e., at the point where half the people in each cohort are alive and half the people are dead) assigns a "life expectancy at birth" value to each cohort. "Life expectancy at birth" is considered a cohort's mean age at death and is used as a basis for statistical analysis and comparison. Thus, the 42 and 47 years life expectancies at birth for the 1850 and 1900 cohorts are significantly greater than the 24-year life expectancy at birth for the 1754 cohort, and the 75 years life expectancy at birth for the 1988 cohort is significantly greater than the life expectancies at birth for the earlier cohorts.
In other words, for nearly two and a half centuries, mean life expectancies in the U.S. and Europe have moved up with statistical regularity. (Life expectancy is higher elsewhere [e.g., Iceland and Japan] and lower elsewhere [conspicuously Africa and Russia].)
But this is the limit of 'squaring'. Indeed, squaring the curve has only a few more years to go before it is squared to saturation! If biopolitics is allowed to continue on its present trajectory, projected life expectancies will increase for white women born in 2100 to 102 years of age, black women and white men to 97, and black men to 90. Even if the conquest of diseases is complete by 2200 as projected by biopoliticians, life expectancy at birth would be 117 years for white women, 112 for black women and white men, and 105 for black men in the US (Olshansky, et al., 1990). This is all that biopolitics has to offer.
The problem for biopolitics arises from the 'entangled tails' as survivorship curves approach 0. This entanglement puts a damper on pushing the curves further outward even with all the power of modern industrialized society lined up behind biopolitics. According to biopoliticians, human beings have a genetically built-in tendency to die sometime before or around 92 years of age.[2] Humans are supposed to hit a biological walla genetic barrierduring the entangled tail phase of the life expectancy curves. According to biopoliticians, during this phase, our probability of surviving from year to year is about 50%. This is not to say that the life of nonagenarians, centenarians, and supercentenarians is necessarily one of decrepitude, but it is a life of chance: the chance of someone sneezing nearby and your catching a cold, flu, or pneumonia that will kill you; of vulnerability to environmental hazards that you would have walked away from earlier in life but now trip you up; and frailty to conditions, like smog, that earlier might have caused annoyance but now threatens to lay you out.
By squaring the curve, the biopoliticians have painted themselves into the proverbial corner. But what about Arakawa and Gins? What does reversible destiny, biotopology, and Bioscleave House have to say about life's limits?
"What limits?"
Biotopology has the potential to extend longevity by disentangling the tails of the survivorship curves. By strengthening the individual, Arakawa and Gins' creation holds the promise of raising the probability of living well beyond a 50% chance.
L. Steven Coles, co-founder of the Los Angeles Gerontology Research Group, created the fifth curve in the illustration (above) by untangling the tails of the survivorship curves and placing the vertical portion's point of inflection at 150 years, but it could be placed virtually anywhere along the continuum. Steve has in mind extending human lifetime by finding ways of expressing salubrious genetic tendencies thereby promoting wellbeing and longevity (personal communication), but he also shares the vision of the creators of Bioscleave House.
Bioscleave House enhances wellbeing by activating and exercising every part of the human organism constantly and productively. By incorporating the contours of a terrain into the contours of an apartment, Bioscleave House flows into a landscape, between rooms, even within rooms, producing an expansive effect in place of the prison cell of four walls and doors. The residents breath more deeply as their horizon expands, exercise their whole body more completely as they move in the interior terrain, and encounter their own artistic spirit as they break away into the "exploratorian" from the quotidian.
But Bioscleave House can also operate on another level by promoting the extension of human longevity through evolution. Once Bioscleave House goes beyond Phase IV and villages of Bioscleave Houses become universal, they will expand life expectancy on the level of the species. Just as Bioscleave House rejects the biopolitical imperative to die, a world of Bioscleave Houses will liberate life from 'squaring the curve.' A world of Bioscleave Houses will open lifetime extension to infinite possibilities.
Arakawa and Gins have shown us how to take control of our destiny and human evolution! It is simply a matter of scale. Enhancing human life will also promote the outward evolution of longevity. Biologists call it "niche construction": how the activities of organisms bring about changes in their environments and, consequently, in their own evolutionhow a species' activity feeds back on the species' environment and hence on its evolution.
Bioscleave House is how we can extend life throughout our species and make it worth living in the process, namely, how we can live longer by living younger! Actually, the process is not new: it is probably responsible for many of the traits that have evolved over the millennia, including our present relatively long life. Juvenilisation, known in the evolutionary literature as "neoteny" (from the Greek meaning stretching, extending or holding onto) refers to the retention of juvenile morphology into adult stages of the lifetime, and hence the delay of aging.
Signs of neoteny are clearly visible in humans Several aspects of the human body strongly remind zoologists of characteristics typical among young, immature, even embryonic forms of primates. Among these are the size of the brain, which is very large in comparison to the rest of the body (like an infant's), the angle of head to spine (a right angle), and a mostly hairless body (Benecke 2002:105). Neoteny is the slowing of somatic development, epitomized by the amphibian mud puppy Necturus maculosus, which retains its larval appearance throughout adult life. But neoteny also occurs widely in other vertebrates, fish, birds, and mammals, andnotablyin humans.
Slow growth is reflected in the delayed age of puberty in women compared to other mammals. [3] Moreover, women experiencing a delay in reaching menopause have not only grown old more slowly than other women but they tend to be longer-lived (Perls et al., 1997). Our aging is also slow compared to aging in other primates. The baboon mortality rate doubles every four years compared to seven to eight years for humans. "[Thus, h]umans. . . age differently, and more slowly than baboons" (Tatar et al. 2009). The pioneering primatologist Sherwood Washburn insists "there is strong direct evidence for the slowing of [human] development" (Washburn 1981: 23).
Furthermore, "[w]hat characterizes modern humans as unique is a prolongation of the postnatal growth period" (Dean 1987: 213). Indeed,
[t]he ages derived for Australopithecus, Paranthropus, and early Homo described biological equivalence to modern man at roughly two-thirds the chronological age, demonstrating that they had growth periods similar to the modern great apes. (Bromage and Dean 1985: 526)
At the end of growth, the adult skull in humans reaches an allometric shape (size-related shape) which is equivalent to that of juvenile chimpanzees with no permanent teeth. (Penin et al. 2002: 50)
Neoteny has other effects: it extends the benefits of juvenile life into adult stages. Juvenilised human beings are healthier, more active, livelier, and more receptive to new ideas than other members of the species. Indeed, one is hardly surprised when the biographers of the French supercentenarian Jeanne Calment describe her at 120 years as "someone who remains very young in spirit, and tastes, a kind of kid, almost childlike at times" (Allard et al. 1998: 62).
And Bioscleave House will only be the tip of the evolutionary iceberg by promoting neoteny and pushing juvenile wellbeing into adulthood. We have yet to conceive of where Bioscleave House will take us by returning us to the sand box of youthful life where life is play, sex is fun, commodities do no harm, creativity expands without leaving waste and where poetry thrives without breeding despair! That is where niche construction will create our future in the here-and-now! Genes will be reshuffled over generations and selection will favor a new, youthful, long-lived Homo sapiens. We will evolve into a species of individuals living younger, living longer and enjoying life all the more. Biotopologistsscientists, poets, artists, architectswill thrive in their Bioscleave Houses forging ahead into appropriate niche construction for reversible destiny, enhanced neoteny, and the evolution of youthful longevity for a lifetime!
Notes
[1] Regrettably, not everyone is doing as well. In fact, 25% of global deaths are still due to infectious diseases striking disproportionately at the young. Even in the United States, the young may not have access to adequate health care.
[2] The fact that Jeanne Louise Calment made it to 122 years and 164 days (born February 21, 1875; died August 4, 1997), surviving two standard deviations beyond the mean for her cohort (a highly significant difference) is simply dismissed as a statistical fluke.
[3] "Human beings reach puberty at an age (1214 years) that is [relatively] 75-fold later than in mice" (Finch 1990: 629).
Bibliography
Allard, Michel, Victor Lèbre and Jean Marie Robine. Jeanne Calment: From Van Gogh's Time to Ours, 122 Extraordinary Years. Trans. Beth Coupland. New York: W. H. Freedman, 1998.
Benecke, Mark. The Dream of Eternal Life: Biomedicine, Aging, and Immortality. Trans. Rachel Rubenstein. New York: Columbia University Press, 2002.
Bromage, Timothy G. and M. Christopher Dean. "Re-evaluation of the Age at Death of Immature Fossil Hominids." Nature 317 (1985): 525527.
Dean, M. Christopher. "Of Faster Brains and Bigger Teeth." Nature 330 (1987): 213.
Finch, Caleb E. Longevity, Senescence, and the Genome. Chicago: University of Chicago Press, 1990.
Foucault, Michel. The History of Sexuality: Vol. 1: An Introduction. Trans. Robert Hurley. New York: Vintage Books, 1980; 1976.
Olshansky, Stuart J., Bruce A. Carnes and C. Cassel. "In Search of Methuselah: Estimating the Upper Limits to Human Longevity." Science 250 (1990): 634640.
Penin, Xavier, Christine Berge, and Michel Baylac. "Ontogenetic Study of the Skull in Modern Humans and the Common Chimpanzees: Neotenic Hypothesis Reconsidered with a Tridimensional Procrustes Analysis." American Journal of Physical Anthropology 118 (2002): 5062. [doi: 10.1002/ajpa.10044]
Perls, Thomas T., Laura Alpert and Ruth C. Fretts. "Middle-aged Mothers Live Longer." Nature 389 (1997): 133.
Tatar, Marc, Andrzej Bartke and Adam Antebi. "The Endocrine Regulation of Aging by Insulin-like Signals." Science 299 (2003), 13461351.
Turner, S.J. "Marc Tartar on the Components of Aging." George Street Journal (2004). file:///research/Frailty/ frailtyMarcTatar 04-13-04_files/redir.html.
Washburn, S. L. "Longevity in Primates." In Aging: Biology and Behavior. Eds. James L. McGaugh and Sara B. Kiesler . New York: Academic Press, 1981. 1129.
Arakawa + Gins
Edited by Jondi Keane & Trish Glazebrook
Open Letters
Madeline Gins i-viii
Here Where it Lives...Biocleave
Jondi Keane and Trish Glazebrook 1-21
NODE:
Mapping Reversible Destiny
Trish Glazebrook and Sarah Conrad 22-40
Escaping the Museum
David Kolb 41-71
Ing
Jean-Jacques Lecercle 72-79
The Reversible Eschatology of Arakawa and Gins
Russell Hughes 80-102
Chaos, Autopoiesis and/or Leonardo da Vinci/Arakawa
Hideo Kawamoto 103–111
Daddy, Why do Things have Outlines?: Constructing the Architectural Body
Helene Frichot 112–124
Tentatively Constructing Images: The Dynamism of Piet Mondrian's Paintings
Troy Rhoades 125–153
Evidence: Architectural Body by Accident, Destiny Reversed by Design
Blair Solovy 154-168
Breathing the Walls
James Cunningham 169–188
Technology and the Body Public
Stephen Read 189-213
Bioscleave: Shaping our Biological Niches
Stanley Shostak 214-224
Arakawa and Gins: The Organism-Person-Environment Process
Eugene Gendlin 225-236
An Arakawa and Gins Experimental Teaching Space – A Feasibility Study
Jondi Keane 237–252
KEYNOTES AND CONFERENCE STREAMS:
The Mechanism of Meaning: A Pedagogical Skecthbook
Gordon Bearn 253–269
Wayfinding through Landing Sites and Architectural Bodies: Exploring the Roles of Trajectoriness, Affectivatoriness, and Imaging Along
Reuben Baron 270-285
Trajectory of ARAKAWA Shusaku: from Kan-Oké (Coffin) to the Reversible Destiny Lofts
Fumi Tsukahara 286-297
A Snailspace
Tom Conley 298–316
Made/line Gins or Arakawa in
Trans-e-lation
Marie Dominique Garnier 317–339
The Dance of Attention
Erin Manning 340–367
What Counts as Language in a Closely Argued Built-Discourse?
Gregg Lambert 368-380
Constructing Poiesis: Storyboards for an immersive diagramming
Alan Prohm 381–415
Open Wide, Come Inside: Laughter, Composure and Architectural Play
Pia Ednie-Brown 416–427
TRIBUTES:
What Arakawa Did
Don Byrd 428–441
Arakawa
Don Ihde 442-445
For Arakawa, Nine More Lives
Jean-Michel Rabaté 446–448
TANGENTS:
Approximately Arakawa and Gins
Ken Wark 448-449
A Perspective of the Universe
Erin Manning and Brian Massumi
450-458
Axial Lecture on Self-Orientation
George Quasha
Demonstrator
Bob Bowen
Levitation
Bob Bowen